
- Home
- Search
- Images
- Species Checklists
- US States: O-Z >
- US National Parks
- Central America
- South America
- US National Parks
- Southern Subpolar Region
|
Rinodina intermedia Bagl.
 |
|
|
Family: Physciaceae
[Rinodina darrowii E. D. Rudolph, moreRinodina sabulosa Tuck.] 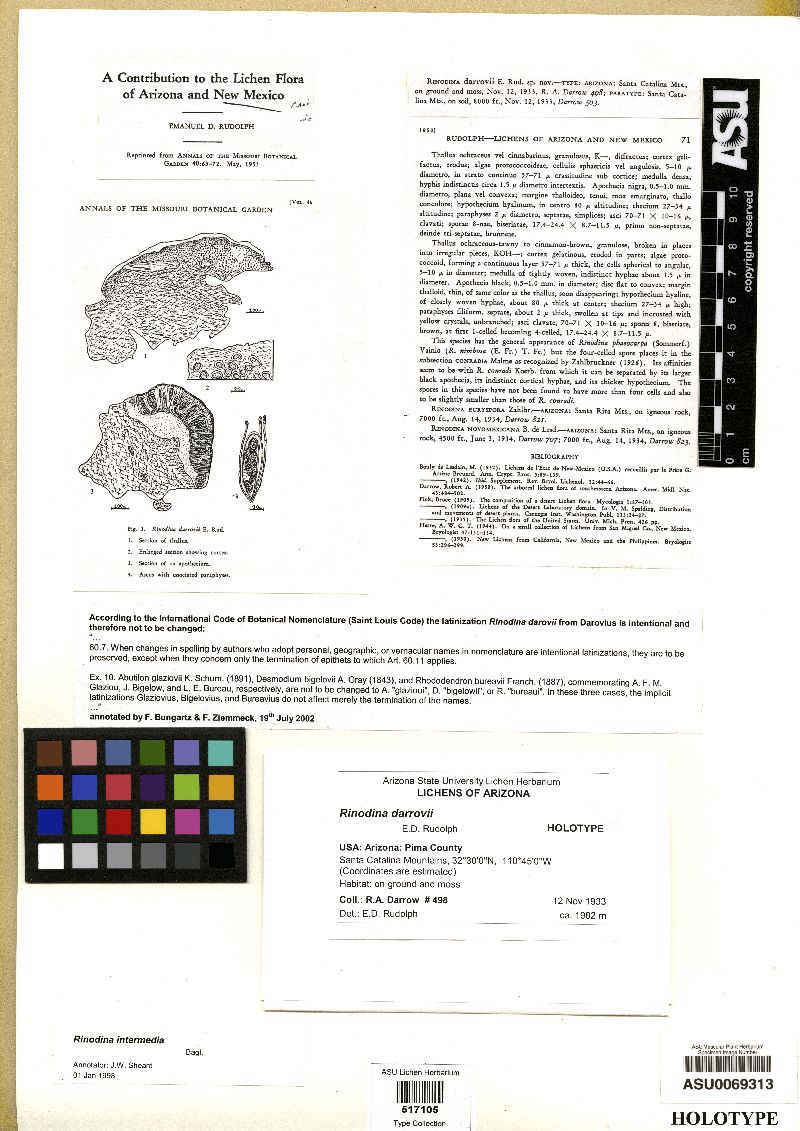 |
MB#404376 Type. [SWITZERLAND.] Sulle rupi conferte da leg. strato di muschi alla Madonna del Sasso, Locarno, Aug. 1862, F. Baglietto s.n. (MOD - holotype). Rinodina sabulosa Tuck.., Lich. California: 21. 1866. Rinodina darrowii E.D. Rudolph [as 'darrovii'], Ann. Missouri Bot. Gard. 40: 69. 1953. Exsiccata. Erbario Critt. Ital. I: 188 (B, M as R. turfacea); II: 514 (BM, G, LD, M, MOD, WU as R. turfacea); Hasse Lich. Exs. 249 (ABS as R. turfacea); Rabenhorst Lich. Eur. 381 (LD, M, MOD as Psora turfacea); Reliq. Tuck. 87 (BM, BP, C, FH, L, LD, M, S, US as R. sabulosa); Sampaio Lich. Port. 199 (LD, LISU, M, US). Description. Thallus thick, thin, or inconspicuous, light grey‑green or ochraceous to light brown, usually continuous, frequently with small marginal lobes, 0.20-0.30 mm wide; surface plane or rugose, matt or shining; margin usually determinate; prothallus lacking; vegetative propagules absent. Apothecia broadly to narrowly attached, frequent and often contiguous, to 0.50-1.10 mm in diam.; disc dark brown to black, plane becoming convex; thalline margin concolourous with thallus, 0.10-0.15 mm wide, entire and persistent, or rarely poorly developed; excipular ring often present, confluent. Apothecial Anatomy. Thalline exciple 70-120 µm wide laterally; cortex 5-10 mm wide, epinecral layer 5-10 mm wide; crystals absent in cortex and medulla; cortical cells to 4.0-6.5 mm wide, pigmented or not; algal cells to 8.5-17.5 mm long; thalline exciple (60-)90-140 mm deep below; cortex 5-30 mm wide; epinecral layer absent or 5-10 (-20) mm deep; proper exciple hyaline, 10-20 mm wide laterally, expanding to 30-80 mm at periphery; hypothecium hyaline, 90-120 mm deep; hymenium 100-140 mm high, not inspersed; paraphyses 2.0-3.0 mm wide, sometimes conglutinate, with apices to 3.0-5.0 mm wide, lightly pigmented and immersed in dispersed pigment to form red-brown epihymenium; asci 80-110 x 19-26 mm. Ascospores 8/ascus, Type A development, 2-4 cells at first, then submuriform, (4-)6-11(-12) celled, Type A development, (20.5-)25.5-27.0(-31.5) x (10.0-)12.0-13.0(-15.0) mm, lumina irregularly rounded; torus absent; walls not ornamented. Pycnidia pear shaped, immersed; conidia bacilliform, 3.5-4.0 x ca. 1.0 mm (Mayrhofer et al. 2001, Figure 6). Chemistry. Spot tests, K- or K+ rose-violet, C-, KC-, P-; secondary metabolites, ± skyrin, deoxylichesterinic acid present in all specimens tested, graciliformin, pannarin, both minor constituents (Mayrhofer et al. 2001). Substrate and Ecology. Rinodina intermedia typically grows on soil or terricolous mosses, rarely on rock, and is often found in shaded crevices between rocks, suggesting that it requires more moisture than is generally available in some of the xeric habitats it frequents. It may rarely occur on the horizontal stems of dead shrubs on the ground, an extension of its usual terricolous habitat. Distribution. In North America R. intermedia has a Sonoran centre of distribution extending as far north as San Francisco (near sea level), California and Larimer Co., Colorado, and to elevations 2,250 m in Chihuahua, Mexico. Widely distributed in dry, warm temperate regions across low latitudes of the northern hemisphere, including the Caribbean, Macaronesia, southern Europe and north to the Channel Islands, U.K., eastwards to the Himalaya and as far south as the equator in Ecuador, South America and Kenya, Africa (Mayrhofer et al. 2001). Notes. Rinodina intermedia is characterized by its 3-septate to typically submuriform spores with type-A development and by its unique chemistry in the genus. It can only be confused with R. conradii Körb. which differs in possessing Type B spore development, strictly 3-septate spores with rhombic lumina and a north temperate distribution. The two species are mainly allopatric. In North America R. conradii extends as far south as coastal Oregon in the Pacific Northwest. Both species occur in Colorado but R. conradii is limited to altitudes over 2,800 m whereas the highest recorded altitude for R. intermedia in the state is 1,770 m. The two species are further compared and discussed by Mayrhofer et al. (2001). The world distribution of R. intermedia is reminiscent of the species discussed by Kärnefelt (1980) with disjunctions between western North America and Macaronesia and the west Mediterranean region, except that it is more extensive being found in the Himalaya, Africa and South America in addition. Kärnefelt suggested that his distributions dated from the Paleogene (66-24Ma). The wider distribution of R. intermedia may indicate an earlier origin before or during the opening of the mid-Atlantic and the beginning of the breakup of Pangea in the Late Jurassic (163-144Ma). The origin of the Himalayan population is of particular interest. Is it a remnant Tethyan population or did it raft north with the Indian subcontinent starting in the Late Cretaceous (97-66Ma)? Specimens examined [not listed by Mayrhofer et al. (2001)]. MEXICO. BAJA CALIFORNIA NORTE. W Cerro Kenton, 1987, T.H. Nash; Guadalupe Island, T.H. Nash 38422; BAJA CALIFORNIA SUR. Cerro Blanco, T.H. Nash 39671. CHICHAHUA, 30 km W Madera, T.H. Nash 36735; Park Nat. Basaseachic, T.H. Nash 37677; W Casa de Junos, T.H. Nash 36119. SONORA, Agua‑Nacozari Road, T.H. Nash 12403; 7 km W Huasabas, J. Marsh 5903; 7 km NE Huasabas, T.H. Nash 14761; Rancho el Encino, T.H. Nash 33565 (all ASU); 5 km W Moctezuma, 1993, M. Krenn (GZU); 10 mi E Nacozari, T.H. Nash 12495 (ASU). U.S.A. ARIZONA. Cochise Co., Carr Peak, T.H. Nash 14548; Pole Bridge Canyon, 1979, L. Sigal; T.H. Nash 15778; Rucker Canyon, B.D. Ryan 10936; Pima Co., Mader Canyon, B.D. Ryan 20324; Santa Catalina Mountains, R.A. Darrow 498, 503 (all ASU); 1933, R.A. Darrow (WIS); Tucson Mountain, C.M. Wetmore 55280, 55501 (MIN); Santa Cruz Co., Sycamore Canyon, T.H. Nash 33524 (ASU). CALIFORNIA. 1865, H.N. Bolander (BM, FH, UC, US, WIS); Los Angeles Co., Casa Verdugo Mountain 1906, H.E. Hasse (FH); Catalina Island, C.C. Bratt 11511 (SBBG); 1895, H.E. Hasse (NY); T.H. Nash 32150, 32116 (ASU); W.A. Weber 42794, 42813 (COLO); Hollywood, 1904, H.E. Hasse (FH); San Clemente Island, W.A. Weber 42672 (COLO); C.C. Bratt 9714, 9856b (SBBG); San Gabriel Canyon, 1902, H.E. Hasse (FH); Santa Monica Mountains, 1897, H.E. Hasse (FH, H, ABSL); 1913, H.E. Hasse (US); H.E. Hasse 258 (MIN, NY, US); Topanga Canyon, 1906, H.E. Hasse (FH); Riverside Co., Chino Canyon, C.C. Bratt 6065 (SBBG); San Diego Co., Camp Kearney, H.E. Parks 117 (COLO, UC); Mesa, 1883, C.R. Orcutt (BM); Point Loma, 1909, H.E. Hasse (FH); R.E. Riefner 9122 (IRVC); C.C. Bratt 8669a, 8670 (SBBG); San Diego, D. Cleveland (US); 1883, C.R. Orcutt (FH); San Francisco Co., Mission Dolores, 1865, H.N. Bolander (PH); H.N. Bolander 76 (US); San Luis Obispo Co., Montana de Oro State Park, P. van den Boom 29185 (personal herb.); Santa Barbara Co., Santa Cruz Island, 1994, S. Sharnoff (CANL); J.W. Sheard 5120 (SASK). COLORADO. Larimer Co., Fort Collins, R.A. Anderson 20320. NEW MEXICO. Union Co., Folsom, S. Shushan 11411. TEXAS. Brewster Co., Chisos Mountains, R.A. Anderson 18791; El Paso Co., N. Franklin Mountain, R.D. Worthington 20992 (all COLO). References. Sheard & Mayrhofer (2002 Fig. 14), Sheard (2004). Nash, T.H., Ryan, B.D., Gries, C., Bungartz, F., (eds.) 2004. Lichen Flora of the Greater Sonoran Desert Region. Vol 2. Thallus: crustose, thick, thin, or evanescent, usually continuous, frequently with small marginal lobes 0.2-0.3 mm wide, plane or rugose surface: light gray-green or ochraceous to light brown, dull or shiny; margin: usually determinate; prothallus: lacking; vegetative propagules: absent Apothecia: adnate to sessile, frequent and often contiguous, up to 0.5-1.1 mm in diam. disc: dark brown to black, plane becoming convex thalline margin: concolorous with thallus, 0.1-0.15 mm wide, entire and persistent, or rarely poorly developed; excipular ring: often present, confluent thalline exciple: 70-120 µm wide laterally; cortex: 5-10 µm wide; epinecral layer: 5-10 µm wide; cortical cells: up to 4-6.5 µm wide, pigmented or not; algal cells: up to 8.5-17.5 µm in diam.; thalline exciple: (60-)90-140 µm thick below; cortex: 5-30 µm wide; epinecral layer: absent or 5-10(-20) µm thick proper exciple: hyaline, 1020 µm wide laterally, expanding to 30-80 µm at periphery hymenium: 100-140 µm tall; paraphyses: 2-3 µm wide, sometimes conglutinate, with apices up to 3-5 µm wide, lightly pigmented and immersed in dispersed pigment to form red-brown epihymenium; hypothecium: hyaline, 90120 µm thick asci: clavate, 80-110 x 19-26 µm, 8-spored ascospores: brown, with 2-4 cells at first, then pseudomuriform, ellipsoid, (4-)6-11(-12) celled, type A development, (20.5-)25.5-27(-31.5) x (10-)12-13(-15) µm, lumina irregularly rounded; torus: absent; walls: not ornamented Pycnidia: pear-shaped, immersed conidia: bacilliform, 3.5-4 x c. 1 µm Spot tests: K- or K+ rose-violet, C-, KC-, P- Secondary metabolites: ± skyrin, deoxylichesterinic acid present in all specimens, graciliformin, ± pannarin (both minor). Substrate and ecology: typically on soil or terricolous mosses and often in shaded crevices between rocks, suggesting that it requires more moisture than is typical for its xeric habitats, also rarely occurring on the horizontal stems of dead shrubs on the ground, an extension of its usual terricolous habitat World distribution: widely distributed in dry, warm temperate regions across low latitudes of the Northern Hemisphere, Macronesia, southern and western Europe and eastwards to the Himalaya and as far south as the equator in South America (Ecuador) and Africa (Kenya), and southwestern North America Sonoran distribution: common in southern California and parts of southern Arizona, southwards into Chihuahua. Notes: Rinodina intermedia is characterized by its 3septate or typically pseudomuriform spores with type-A development and by its unique chemistry in the genus. It can only be confused with R. conradii Körb. that differs in possessing type-B spore development and strictly 3-septate spores. Rinodina conradii does not occur in the Sonoran region, having a cool temperate distribution. The two species are discussed at length by Mayrhofer et al. (2001). It is the only species with a Sonoran distribution in the sense of McGlaughlin (1989). |

")


")


")


")




")

")


")
")
![]()
![]()

![]()
![]()
![]()
![]()
![]()