
Consortium of Lichen Herbaria
- building a Global Consortium of Bryophytes and Lichens as keystones of cryptobiotic communities -
- Home
- Search
- Images
- Species Checklists
- US States: O-Z >
- US National Parks
- Central America
- South America
- US National Parks
- Southern Subpolar Region
|
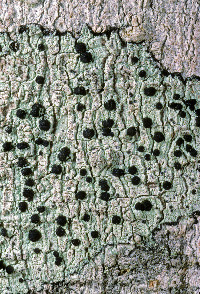
Buellia erubescens Arnold
 |
|
|
Family: Caliciaceae
Common Button Lichen
[Buellia stillingiana J. Steiner, moreBuellia zahlbruckneri J. Steiner, Buellia zahlbruckneri var. erubescens J. Steiner, Buellia zahlbruckneri var. zahlbruckneri J. Steiner]  Stephen Sharnoff |
Nash, T.H., Ryan, B.D., Gries, C., Bungartz, F., (eds.) 2007. Lichen Flora of the Greater Sonoran Desert Region. Vol 3. Lectotype: selected here for B. stillingiana: U.S.A. Tennessee: Hamilton Co. Lookout Mountain, Calkins 136 (!MICH-0063535) Thallus: crustose, rimose to rimose-areolate, thin to moderately thickened, ±continuous; prothallus: absent or delimiting the thallus as a black outline where several different thalli meet surface: gray, grayish white to ivory, dull, smooth, epruinose, phenocorticate, esorediate medulla: white, lacking calcium oxalate (H2SO4-) Apothecia: lecideine; (0.2-)0.3-0.7(-0.9) mm in diam., soon sessile margin: black, thick, rarely persistent, usually excluded with age, rarely with a thalline veil when emerging from the thallus disc: black, epruinose, plane, soon becoming convex proper exciple: dispersa-type, inner excipular hyphae distinct, not reduced, pigmented, prosoplectenchymatous (textura oblita), extending from the deep reddish brown hypothecium (leptoclinoides-brown, textura intricata), outer excipular hyphae short-celled, cells angular, distinctly swollen (textura angularis) and ±carbonized with various amounts of a brown pigment (cf. elachista-brown, HNO3-) epihymenium: brown, pigmentation continuous with the outer exciple (HNO3-) hymenium: hyaline, not inspersed with oil droplets; paraphyses: simple to moderately branched, apically swollen, with a brown pigment cap (cf. elachista-brown) asci: clavate, Bacidia-type, 8-spored ascospores: soon brown, 1-septate, ellipsoid to broadly ellipsoid, not constricted, with obtuse ends, not curved, (9-)11.3-[13.1]-14.9(-16) x (6.5-)6.7-[7.3]-7.9(-8) µm (n=26); proper septum: soon but only very briefly thickened, lateral wall never thickened (Buellia-or indistinct Physconia-type); ornamentation: very faintly microrugulate (best seen in DIC) Pycnidia: rare, urceolate to globose, unilocular; ontogeny similar to the Umbilicariatype conidiogenous cells: mostly terminal, rarely also intercalary (cf. conidiophore-type V) conidia: bacilliform, 6-9 x 0.5-1 µm (n=29) Spot tests: K+ yellow to orange red (sometimes forming crystals), P- to weakly yellow, C- fluorescence: UV-(pale) iodine reaction: medulla non-amyloid Secondary metabolites: chemotype I: atranorin, norstictic acid usually accompanied by connorstictic acid, traces of salazinic, hyposalazinic acid; chemotype II: atranorin, ±traces of norstictic and connorstictic acid. According to Giralt et al. (2000) specimens may also contain ±zeorine and an unknown secalonic acid derivate(s). Material from the Sonoran region typically contains only small amounts of norstictic acid. Small amounts of usnic acid and fulgidin were also detected in Sonoran specimens (J. A. Elix, HPLC). Substrate and ecology: on bark or rarely wood of Quercus, Pinus, Juniperus, or trees with bark of generally low pH, common in submontane to subalpine forests (600-3000 m) World distribution: widely distributed throughout the USA and Europe Sonoran distribution: at inland localities of Arizona and Sonora, and also in southern California and Baja California Sur at lower altitudes. Notes: Imshaug (1951) did not discuss B. erubescens, and apparently was unaware of that taxon. He, however, did distinguish B. zahlbruckneri from B. stillingiana. Buellia stillingiana is, however, a synonym of B. erubescens, a fact not previously recognized because of confusion of B. erubescens with B. zahlbruckneri (Giralt et al. 2000). Buellia zahlbruckneri sensu Imshaug (1951) is not a synonym of B. erubescens, even though the type material of B. zahlbruckneri, as discussed by Giralt et al. (2000), clearly belongs to that species. In his discussion Imshaug (1951) compared B. zahlbruckneri with B. parasema (cf. B. disciformis), citing oil inspersion in the hymenium as the main difference for the two taxa. Furthermore, Imshaug (1951) reported that spores of B. zahlbruckneri are ovate-oblong, occasionally slightly constricted. Most specimens identified as B. zahlbruckneri by Imshaug accordingly belong to B. chloroleuca, a species with little or no oil inspersion, slender and sometimes constricted spores, and with xanthones. In high concentrations these xanthones are easily recognized by their C+ orange spot test reaction, a test that Imshaug (1951) did not report. As one of the most important characters to recognize B. stillingiana, Imshaug (1951, p. 116) emphasized a ?? Yellowish or pale yellow-brown parenchymatous tissue which extends into the thallus??. Brodo (1968, p. 144) subsequently regarded this yellow hymenium as one of the most diagnostic characters. However, in the type material of both B. erubescens and B. stillingiana, a yellowish pigment is present throughout the hypothecium, not the hymenium. If the hypothecial pigment extends from the hypothecium towards the hymenium, a yellowish subhymenium might be distinguished between hymenium and hypothecium. In other specimens where it does not extend into the subhymenium, the pigment may be difficult to locate against a generally deep brownish red hypothecium. The distinctiveness of the pigmentation also depends on the thickness of apothecial sections that are studied. In thin sections pigment concentration might be very low and thus easily overlooked. In thick hand sections, almost the entire hymenium often appears to be pigmented. Because of the variation of this yellow pigmentation, we are not convinced that the character is as reliable for species identification as previously suggested. Although the pigment intensifies somewhat in K, this reaction may also be attributed to the presence of norstictic acid as well as atranorin, as both substances are typically present at least as traces. The obscuring reaction of the secondary metabolites hinders the identification of the yellow pigment according to color reactions sensu Meyer and Printzen (2000). According to Giralt et al. (2000), spores of B. erubescens are roughly ornamented?. This is not the case according to our own studies, which included type material of B. erubescens, B. zahlbruckneri, and B. stillingiana. All specimens have smooth to very faintly microrugulate spores. In the Sonoran Region, B. erubescens can reliably be distinguished from superficially similar species like B. chloroleuca, and B. disciformis by its ascospores, which are not constricted at the septum, smooth, and ellipsoid to broadly ellipsoid. Young ascospores are typically fairly broad and show a ±distinct septal thickening, which, however, soon disappears. Mature ascospores have a narrow septum and are slightly more elongate, but never taper as obviously as spores of the previously mentioned species. Spores of B. erubescens never show any tendencies to develop lateral spore wall thickenings, which are characteristic for the Callispora-type present in the other species. Reports of Amandinea submontana Marbach from the Sonoran Region are based on misidentifications of specimens that either belong to B. erubescens or to B. chloroleuca. |
 Stephen Sharnoff  Stephen Sharnoff  Stephen Sharnoff  Stephen Sharnoff  Stephen Sharnoff  Stephen Sharnoff  Stephen Sharnoff  Stephen Sharnoff  Lucy Taylor  Sheila Strawn  Gary Perlmutter  ") ")  ") ")  Andrew Khitsun  Andrew Khitsun  Andrew Khitsun  Gary Perlmutter  Gary Perlmutter  Gary Perlmutter  Gary Perlmutter  Gary Perlmutter  Gary Perlmutter 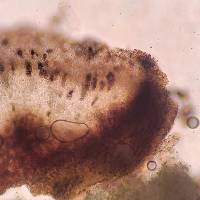 Gary Perlmutter  Gary Perlmutter 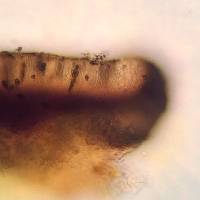 Gary Perlmutter  Gary Perlmutter  Gary Perlmutter ") ") ") ") ")   ")  ") ")  ")  ")  ")  ") ") ")   ")   ") Christopher Raithel ")   ")   ") ") ") ") ")  Gary Perlmutter ") ") ")    Jason Hollinger ") ") ") ") ") ") ")  ")  ") ") |
|

![]()
![]()

![]()
![]()
![]()
![]()
![]()
Powered by Symbiota